
Tutte le cellule del nostro corpo sono delimitate da una membrana che è costituita da un doppio strato fosfolipidico costituito da teste idrofile e code idrofobiche. Tale membrana è di fondamentale importanza nella determinazione delle proprietà elettriche della cellula e dell’eccitazione cellulare. Lo scopo della membrana è quello di separare soluzioni acquose che presentano, disciolti al loro interno, atomi di sostanze diverse tra loro.
La separazione è utile ai fini dell'assorbimento selettivo da parte della cellula dei soli composti e “ioni” necessari al suo funzionamento. Ai capi della membrana, quindi, si vengono a creare delle differenze di potenziale dovute alla concentrazione degli ioni. Tra gli altri troviamo Potassio (K), Sodio (Na) e Cloro (Cl), la cui concentrazione dimostra un gradiente. All'esterno della cellula, vi è, infatti, una maggiore concentrazione di ioni Na+ mentre all’interno della membrana cellulare vi è una più alta concentrazione di ioni K+ (il rapporto è di circa tre a uno). Questa disposizione è una diretta conseguenza della impermeabilità della membrana cellulare, ovvero della difficoltà con la quale può avvenire l'attraversamento diretto della stessa da parte di uno ione; questo passaggio è, infatti, permesso solo attraverso apposite strutture trans-membranose dette canali ionici, altamente specifiche e che possono essere presenti in conformazione aperta o chiusa, permettendo o meno il transito degli ioni secondo il loro gradiente.
La concentrazione degli ioni potassio è tanto maggiore all’interno della cellula nervosa quanto essa ha un potenziale a riposo tale che la sua permeabilità al potassio è 50-100 volte maggiore rispetto al sodio. Ne discende che il potassio diffonde con molta più facilità del sodio, in quanto a riposo vi sono numerosi canali, specifici per il potassio, che si trovano nella conformazione aperta, permettendo il flusso di ioni. Esistono meccanismi di attraversamento della membrana che possono svolgersi sia secondo sia contro gradiente. Risulta evidente che questo secondo caso non può verificarsi spontaneamente ed è necessario che sia consumata una certa quantità di energia.
Un'altra conseguenza dell’entità dei valori delle concentrazioni è la formazione del cosiddetto potenziale di membrana.
In questa situazione, infatti, il ruolo giocato dalla membrana cellulare appare molto simile a quello del dielettrico in un condensatore a piani paralleli. Così come accade in un condensatore, anche qui sono possibili fenomeni di scarica che abbassano repentinamente l'entità dei gradienti di concentrazione come avviene, per esempio, nella trasmissione di impulsi nervosi.
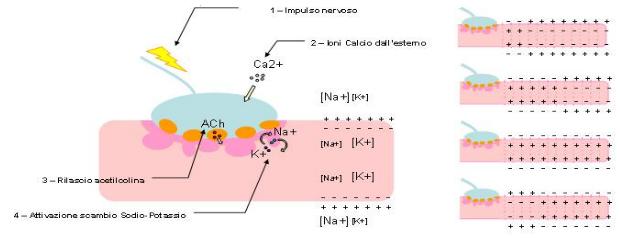
La struttura all'interno della quale questo fenomeno si verifica è costituita dalla terminazione nervosa che termina in una serie di espansioni bottoniformi accolte in invaginazioni della fibra muscolare (come evidente in Figura 1.6). La membrana dell’assone resta separata da quella della fibra muscolare da uno spazio di 20-50 nm. Le fibre muscolari nel corpo umano possono essere di due tipi: striate o lisce. Dal punto di vista funzionale, ciò che li differenzia è il tipo di stimolazione a cui sono sensibili (stimoli volontari, nel caso di muscolo striato; stimoli involontari, nel caso di muscolo liscio). Ciò che avviene in entrambi i casi, però, è analogo.
Il fenomeno dell'impulso nervoso è dovuto all'accumulo di un neurotrasmettitore, l'acetilcolina (ACh, C7H16NO2), proprio nella regione compresa tra le due membrane.
L'eccesso di questa sostanza è responsabile della variazione della permeabilità della membrana (perché legando il proprio recettore, che costituisce un canale per il sodio, lo apre) ed è, dunque, proprio questo che causa l'attraversamento massivo della stessa da parte degli ioni sodio. In condizioni di riposo, l'acetilcolina è comunque sempre presente ma non ve n’è un eccesso insieme ad essa è presente anche la noradrenalina (NE, C8H11NO3) che ha, qualora non sia necessario trasmettere alcuno stimolo, la funzione di contrastare l’acetilcolina ed inibire l’aumento di concentrazione. L’acetilcolina è conservata in massima parte in vescicole all’interno dei neuroni; nel momento in cui c’è necessità di una contrazione muscolare, cioè quando vi è uno stimolo, essa viene rilasciata.
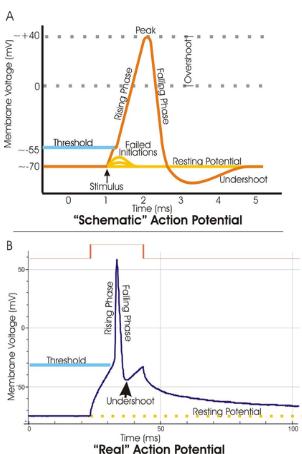
Come conseguenza di questo tipo di stimoli, la tensione transmembranosa subisce delle variazioni al termine delle quali essa torna a valori di riposo. Il grafico in Figura 1.7 dimostra l'andamento del potenziale. Si vede chiaramente, infatti, che la tensione ai capi della membrana (indicata in millivolt) si attesta intorno a 70 mV a riposo. All'aumentare della differenza di potenziale, ed una volta superato il valore di soglia, indicato intorno a 55 mV, l'effetto dell'attraversamento massivo degli ioni sodio attraverso la membrana causa proprio quella scarica di cui abbiamo discusso che, altro non è, che il vero è proprio impulso nervoso. La fase discendente dimostrata in Figura 1.7 non è altro che l'attraversamento della membrana ad opera degli ioni potassio che ristabilirà il potenziale di riposo iniziale. Come si vede dal grafico, l'intera escursione del fenomeno "impulso" ha una durata di circa 2 ms ed un’escursione di 0,1 V. Una volta a riposo, la pompa sodio-potassio ripristinerà la concentrazione ionica originaria ai capi della membrana. La più bassa corrente necessaria per ottenere l'attivazione cellulare è detta corrente reobasica o reo base. Teoricamente questa corrente necessita di una durata infinitesima per l'avvio. Il tempo necessario per eccitare la cellula con il doppio della corrente reobasica è detto cronassiale.
È piuttosto importante sottolineare che quando avviene l'attivazione cellulare, la membrana è insensibile ad altri stimoli, indipendentemente dalla loro intensità. Questa fase è nota come "periodo refrattario assoluto". Al termine, invece, dell'effetto impulsivo attivazionale, la cellula può essere attivata solamente con uno stimolo più forte del normale e questa fase è nota come "periodo refrattario relativo".
Il meccanismo appena descritto è relativo ad una sola terminazione nervosa. Per comprendere come funzioni l'intero sistema nervoso, indipendentemente dalla volontarietà del soggetto, è necessario immaginarsi un insieme discreto ma denso di terminazioni nervose che si irradiano lungo le fibre muscolari. Questa struttura risulta sensibile non solo a stimoli endogeni ma anche a quelli esterni. L'intensità di una sensazione rilevata quale effetto di uno stimolo esterno è dovuta ad un vero e proprio processo di modulazione di frequenza attuato dal sistema nervoso agli impulsi nervosi trasmessi dacché la frequenza degli impulsi viene modulata sulla base dell’intensità del segnale.
L'indice completo degli articoli relativi alla tesi di laurea sulla interazioni e sugli effetti delle radiazioni sul corpo umano, è disponibile qui



Caro ing. Piero,
hai visto report su rai3 domenica scorsa? Diciamo che successivamente a quella trasmissione si sono avute alcune repliche in altre trasmissioni e dopo di nuovo il silenzio.
Nel prossimo futuro contribuiranno anche le trasmissioni digitali a peggiorare la situazione, perche’ il nocciolo della questione sta nel tipo di modulazione ed e’ quella digitale che e’ dannosa.
Da sempre siamo irradiati dal sole con una certa quantita’ di OEM circa 1kW/mq che non e’ poco, ma non fa male se escludiamo l’eventuale eccesso di UV e oltre soggiornando molto tempo al sole, o gli IR a cui ci sottraiamo perche’ la nostra pelle e’ sensibile ad essi (quando sentiamo caldo al sole, in maniera naturale ci spostiamo all’ombra) a nulla servono le attuali misure di intensita’ di campo EM…
cordialmente
Luigi Papaleo
…non capisco dove sia il problema.
O meglio, non capisco il senso di questo post.
Pare quasi lei mi voglia dimostrare qualcosa… Mi corregga se mi sbaglio ma mi pare come se lei avesse desiderio di sottolineare una mia inesattezza.
È corretto?!
Nella tesi di capi pulsati si parla, mica di campi continui…
Inoltre la trasmissione in questione avevo qualche pecca. È apparsa, sul piano tecnico, piuttosto carente. Tutta la parte dedicata all’analisi della questione dei finanziamenti invece è stata piuttosto stimolante.
Inoltre la sentenza del 31 maggio 2011 dimostra come le conclusioni del mio lavoro, datato luglio 2010, siano valide, attendibili e veritiere.
La conclusione che “a nulla servono le attuali misure di intensita’ di campo EM” risulta, peraltro, parzialmente vera. È giusto, infatti, ripensare a quel modello di analisi e tipizzazione delle misure dosimetriche per poter approfondire e standardizzare dei criteri meno sottomessi al conflitto di interessi che vivono i produttori di cellulari.
Con simpatia.
Piero Boccadoro.